|
|
|
|
ACIDUL RIBONUCLEIC - CARACTERISTICI GENERALE
-structura monocatenara (alcatuita dintr-o singura catena)
Arn-ul este de 3 tipuri ce sunt esentiale pentru realizarea procesului de BIOSINTEZA PROTEICA:
1) arn mesager=copiaza informatia genetica din nucleu
2) arn de transfer=transfera amino-acizii din citoplasma pana la nivelul ribozomilor
3) arn ribozomal=alcatuieste organisme celulare comune la nivelul carora se realizeaza biosinteza proteica.
ARN-ul, spre deosebire de ADN, este o macromolecula alcatuita, de regula, dintr-o singura catena polinucleotidica care se formeaza tot prin legaturile diestericedinte radicalul fosfat si pentoza. Moleculele ARN nu pot avea dimensiuni foarte mari deoarece, cu cat creste numarul nucleotidelor (peste cateva mii ) cu atat stabilitatea moleculei scade.
Sinteza ARN (transcrierea) se realizeaza tot pe baza complementaritatii bazelor azotate ca si in cazul replicatiei ARN. Cele doua catene ale moleculei ADN se despart pe intervalul care urmeaza a fi transcris, numai ca de data aceasta va actiona enzima ARN polimeraza. Acum se va transcrie numai una din catene din molecula ADN:catena sens care va servi ca matrita. Nucleotidele libere care se vor alinia pe baza complementaritatii vor contine riboza. In dreptul adeninei de pe catena veche acum se va atasa uracilul in catena nou sintetizata.
Dupa formarea catenei, molecula ARN paraseste locul transcrierii iar catenele ADN revin la pozitia initiala.
ARN este purtator unic al informatiei ereditare la virusurile ARN(ribovirusur) si la viroizi. Acestia din urma au doar o molecula mica da ARN fara invelis proteic. Ei produc unele boli la plante(boala tuberculilor fusiformi la cartofi).
La restul organismelor, ARN contribuie in diferite moduri la structura si functionarea materialului genetic existand de aceea mai multe tipuri de ARN.
ARN mesager(ARNm) are rolul de a copia informatia genetica dintr-un fragment de ADN si de a o aduce, ca pe un mesaj, la locul sintezei proteice. Moleculele sunt todeauna monocatenare si au lungimi diferite, in functie de marimea moleculelor care urmeaza a fi sintetizate.
ARN ribozomal(ARNr) intra in alcatuirea ribozomilor asociat cu diferite proteine. El este sintetizat tot prin transcrierea din ADN, dupa care catena ARNn se pliaza formand portiuni bicatenare datorita complementaritatii bazelor azotate. Un ribozom este format din doua subunitati care vor recunoaste (tot pe baza complementaritatii) si vor atasa intr ele nucleotidele de recunoastere de la inceputul moleculei de ARNm. Ribozomii au fost descoperiti de savantul George Emil Palade, laureat al premiului Nobel.
ARN de transfer(ARNt) este specializat pentru aducerea aminoacizilor la locul sintezei proteice. Molecula este formata din 70-90 de nucleotide. Are portiuni bicatenare care ii dau forma unei frunze de trifoi. Are doi poli functionali:
a) unul la care se ataseaza un anumit aminoacid;
b) altul care contine o secventa de 3 nucleotide care recunoaste o anumita secventa ARNm unde se ataseaza pe baza complementaritatii.
Alte tipuri de ARN intra in constitutia cromozomilor atat la procariote cat si la eucariote.
Cromozomii
Cromozomul este alcatuit dintr-o macromolecula de ADN, in constructia caruia intra mai multe gene lincate.
Cromozomii indiferent de modul de organizare fizica si moleculara,sunt unitati genetice permanente,care isi conserva individualitatea,proprietatile structural- functionale si se transmit prin auto-aplicare de-a lungul generatiilor celulare.
Exista doua tipuri de cromozomi:
-cromozomul de tip procariot
-cromozomul de tip eucariot.
1) Cromozomul de tip procariot:
-este caracteristic bacteriilor si algelor albastre-verzi
-are o organizare relativ simpla si este format dintr-o macromolecula de ADN,dublu catenara,elicala circular covalent inchisa , cu o varietate de proteine.
GENOMUL VIRAL: este alcatiut dintr-o molecula de ADN (acidul dezoxiribonucleic) sau ARN (acidul ribonucleic),
unucatenara sau bicatenara,liniara sau circulara,inchisa intr-un invelis de proteine.
- are o functionalitate particulara
-la viroizi genomul este in exclusiv ADN
-la eucariote are o organizare mai complexa in sensul ca ADN este permanent compexat cu proteine histonice si alte proteine.
Fiecarei specii ii apartine un anumit numar de cromozomi,de forma si structura bine definite;astfel organismele eucariote pot fi identificate prin complementul lor cromozomal.
Toate celulele din constitutia corpului unui organism pluricelular au aceiasi numar de cromozomi,acesta fiind in numar de doi.
Celulele reproducatoare poseda numai un singur set de cromozomi si se numesc haploide.
NUMARUL DE COMOZOMI DIN CADRUL SETULUI HAPLOID este o constanta pentru fiecare specie.
majoritatea speciilor de plante ,animale si fungi au complemente cromozomale formate din 10-50 de cromozomi.
FORMA cromozomilor metafizici este adesea de betisoare;uneori pot fi filoformi sau granulari.
DIMENSIUNILE cromozomilor sunt in general de ordinul micronilor.
AUTOZOMII sunt cromozomo normali ai complementului unei specii.Ei poarta gene care controleazadiferite caractere;morfo-anatomice si eco-fiziolologice ale organismelor.
HETEROZOMII sunt cromozomi care contin un set de gene cu rol decisiv in determinarea sexului unui individ.
2) Sructura cromozomului de tip eucariot:
Cromozomi sunt structuri cu o morfologie dinamica
-aspectul lor se modifica pregnant in timpul ciclului celular
-cromozomul profazic si metafizic este format din doua elemente fibriliare numite cromatine.
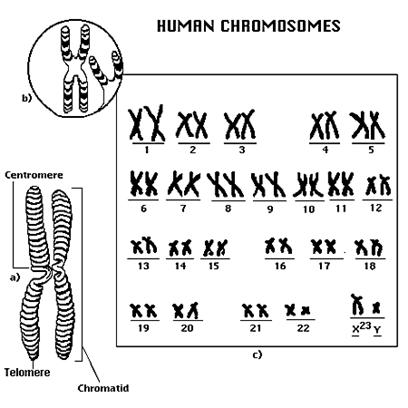
Cele doua cromatine a unui cromozom sunt omoloage din punct de vedere morfologic,biochimic,genetic si functional;una reprezinta copia celeilalte,deoarece rezulta in urma unui fenomen de replicare(copiere)de unde si numele de 'cromatide surori.'
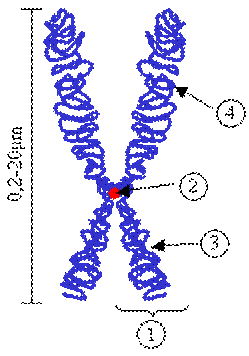
Comadidele sunt unute intre ele printr-o regiune numita centromer.
Centromerul delimiteaza in cadrul unui cromozom doua portiuni egale sau inegale numite brate.
uneori centromerul este situat la unul dintre capetele cromozomului;in acast caz cromozomul are un singur brat.
Dupa pozitia centromerului se disting mai multe tipuri morfologice de cromozomi(metacentric,submetacentric,subtelocentric,telocentric sau acrocentric.La oanumita specie se gasesc in mod constant tipuri morfologice particulare de cromozomo; identificarea acestora este de mare utilitate pentru cunoasterea cariotipului uneu specii, a carui analiza esta deosebit de relevata in caracterizarea acesteia
Prin cariotip se intelege setul de cromozomi caracteristic unei anumite specii.
1=CROMATIDA, 2=CENTROMER,3=BRAT SCURT,4=BRAT LUNG
ORGANIZAREA MATERIALULUI GENETIC LA:
A.VIRUSI
Virusul este o particula submicroscopica, care este formata dintr-o teaca, sau invelis cu rol de protectie de natura proteica, numita capsida si un material genetic. Materialul genetic poate sa fie ADN, fie ARN.
VEZI SI CLASIFICAREA DE LA CLASA A IX A.
B.PROCARIOTE
Materialul genetic la procariote se caracterizeaza printr-o serie de particularitati, si anume:
Cromozomul bacterian este alcatuit dintr-o molecula de ADN dublu catenara circulara. Lungimea ei variaza intre 1000-1400 m, diametrul fiind de 2,5 nm. Numarul de nucleotizi la E.coli este de 4,1·106, iar masa moleculara de circa 2,5· 109 daltoni.
Se presupune, ca nucleoidul vazut la microscopul electronic reprezinta ADN-ul condensat, genetic neactiv (Ryter, Cohen, 1975), iar secventele de ADN ce se transcriu sunt prezentate sub forma de bucle, externe nucleoidului.
Numarul de gene in cromozomul bacterian este de cateva mii (de exemplu, la E.coli - circa 4000).
Dupa functia lor, genele respective se divizeaza in:
Odata cu descoperirea importantei acizilor nucleici in biosinteza proteinelor si a mecanismelor de realizare a mesajului genetic a aparut problema modalitatii de asigurare a cantitatii optime de proteine necesare organismului. Se stie, ca sinteza unei proteine sau a unei enzime nu este constanta in timp, ea trebuie sa se adapteze variatelor cerinte ale functiei celulei.
C.EUCARIOTE
La eucariote materialul genetic are o organizare cu mult mai complexa. Moleculele liniare de ADN, dublu catenare, formeaza complexe cu proteinele histonice, alcatuind fibra elementara nucleohistonica (nucleozomala) (fig.2). Aceste substante, completandu-se cu ARN si proteinele nehistonice, formeaza cromatina, materialul din care se diferentiaza, in procesul de diviziune, cromozomii.
De mentionat ca la eucariote materialul genetic se caracterizeaza printr-o discontinuitate genetica (in molecula de ADN se succed secvente informationale (exoni) si neinformationale (introni), care sunt eliminate in procesul de transcriptie a informatiei genetice (fig.3).
Purtatorii materiali ai informatiei ereditare sunt considerati cromozomii. Termenul a fost propus in anul 1888 de catre W.Waldeyer. Acesta a descoperit niste corpusculi cromatici in celulele in diviziune.
Cromozomii (din gr."chroma" - culoare, "soma" - corp) reprezinta unitati structurale compacte, constante, alcatuite din acizi nucleici si proteine, vizibili in timpul diviziunii celulare la o tratare cu coloranti bazici. Cromozomii au un rol deosebit in viata celulei (organismului), deoarece asigura transmiterea caracterelor la descendenti.
La organismele eucariote, genele nu sunt organizate in operoni, sistemele de reglaj fiind mult mai complicate decat la bacterii. In general, reglarea activitatii genelor la eucariote este mai putin studiata. Aceasta situatie este determinata de o serie de factori, printre care: existenta nucleului separat de citoplasma, structura complicata a cromozomului, influenta sistemelor generale de reglare a organismului, in special a hormonilor etc.
S-a descoperit ca unii hormoni steroizi pot determina activitatea/inactivitatea anumitor gene. Conform datelor lui Watson si col.(1983), acesti hormoni se fixeaza in citoplasma pe moleculele de receptori specifici. Aceste complexe receptor-hormon patrund in nucleu (in timpul interfazei), unde mai apoi se unesc cu cromozomii si afecteaza (pozitiv sau negativ) sinteza proteinelor.
S-a stabilit ca ARNm din celulele eucariote sunt monocistronice, astfel ca un ARNm codifica o singura proteina.
La eucariote, in cromozomul condensat cu histone, ADN-ul nu functioneaza ca matrice pentru transcriptia ARNm, genele sunt in stare de represare. Trecerea acestor gene in stare activa este posibila doar prin inlaturarea histonelor si eliberarea ADN-ului. Eliminarea histonelor are loc sub actiunea histon-proteazei.
Reglarea expresiei genelor la organismele eucariote se face, in primul rand, la nivelul transcriptiei. In unele cazuri, reglarea se poate manifesta si in momentul decodificarii mesajului la nivelul translatiei. Astfel, unele proteine sunt capabile de a se fixa intr-o maniera selectiva pe propriul ARNm, in acest fel impiedicand ribozomii sa sintetizeze proteina respectiva in cantitate mai mare.Spre deosebire de genele procariote, care sunt unitare atat in replicare, cat si in transcriptie si translatie, genele organismelor eucariote sunt unitare in replicare, dar neunitare si discontinue in transcriptie si translatie. S-a demonstrat ca genele organismelor eucariote sunt fragmentate in exoni si introni.Intronii si exonii intr-o gena ocupa pozitii stabile, si sunt variabili in numar si marime. Astfel, gena ce codifica actina (proteina necesara motilitatii celulei) poseda doi exoni si un singur intron. In genele ce codifica fiecare dintre lanturile hemoglobinei, exista trei exoni separati si doi introni. In gena ce codifica colagenul (proteina ce asigura elasticitatea pielii), exista cateva zeci de introni.Cercetarile efectuate de catre Slonimski si col. (1978, 1980) au demonstrat ca intronii participa la reglarea expresiei genelor prin controlul cantitatii de proteine sintetizate sub actiunea genelor.Ipotetic, se considera ca intronii au fost prezervati de evolutie in cromozomii eucariotelor, deoarece ei au un rol particular, prezent doar la eucariote si absent la procariote. Pornind de la diferenta fundamentala dintre procariote si eucariote (prezenta diferentierii celulare la organismele superioare), a aparut ipoteza privind existenta unei legaturi dintre introni si reglarea expresiei genelor.De mentionat ca reglarea activitatii genelor in celulele eucariote are loc pe tot parcursul procesului de realizare a informatiei genetice, de la transcriptie pana la translatie. Deosebim cinci niveluri principale de reglaj genetic:
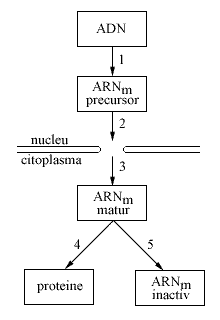
Fig.4. Nivelurile reglajului genetic la eucariote.
Desi reglajul genetic se poate realiza la oricare dintre nivelurile mentionate, cel mai important la eucariote este nivelul transcriptional.
Genomul nuclear este reprezentat de
cromatina-un set de molecule liniare de ADN, fiecare reprezentand un
cromozom. Cromatina este forma
interfazica a cromozomilor care apar individualizati doar in timpul
diviziunii celulare.Numarul de cromozomi este specie specific si nu
are legatura cu complexitatea organismului ,deoarece
marimea,structura si compozitia cromozomilor este foarte diferita.
Cromatina are 2 stari
functionale si alternative: eucromatina si
heterocromatina.Eucromatina are un ciclu de condensare standard -condensare in
diviziune si decondensare in interfaza si contine gene
majore;
Heterocromatina este condensata si in interfaza, e inactiva
in transcriptie, are functii reglatoare.
Un fragment de ADN din 146 nucleotide rasucit de 2 ori pe un octamer de proteine histonice- 2H2A;2H2B; 2H3;2H4. Diametrul = 10 nm
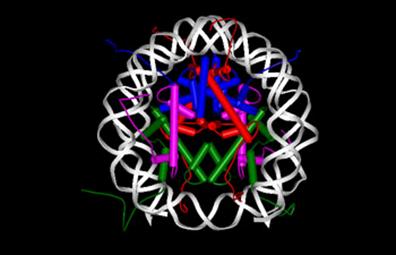
NUCLEOSOMUL
Nucleosomii sunt legati intre ei de un fragment de ADN linker de 140 nucleotide ce e sustinut la mijloc de o proteina histonica H1
Al doilea stadiu de compactare il reprezinta structura de solenoid cu diametrul de 30nmsi 6 nucleosomi/spira.
Aceasta structura e caracteristica in interfaza- fibra de cromatina.
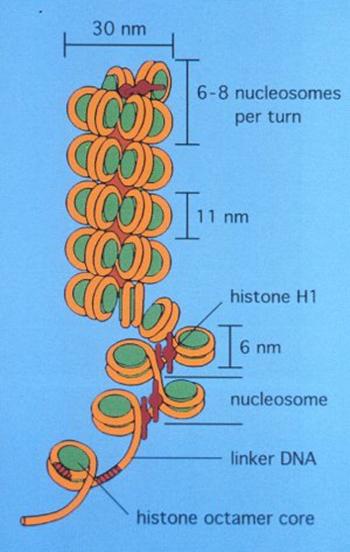
Condensarea maxima a cromatinei este in cromozomii metafazici, in care materialul genetic este condensat de 9000 ori, diametrul e de 700nm si se formeaza cromozomii metafazici.
Structura cromozomilor metafazici
2 cromatide unite
-printr-un centromer
Prin tehnici speciale se pot identifica in cromozomi zone cu condensare diferita a cromatinei
(eucromatina si heterocromatina)- bandarea cromozomilor, cu rol in identificarea cromozomilor si a mutatiilor.
Notiunea de cromozom inseamna in sens strict corpuscul colorat. La microscop cromozomii pot fi evidentiati in cursul diviziunii celulare ca niste formatiuni avand un structura si un numar caracteristic pentru o anumita specie. Ei asigura transmiterea informatiei genetice de la o generatie celulara la alta. Cromozomii fiind caracteristici ca marime, morfologie si numar pentru fiecare specie, constituie un criteriu de identificare taxonomica a acestora.
La inceputul mitozei, cromozomii care au suferit o duplicatie
in cursul interfazei apar la microscop ca fiind formati din doua subunitati
identice denumite cromatide, care sunt unite intr-un singur loc denumit
centromer.Dupa aceea cromatidele se separa complet si devin cromozomii fii,
care migreaza in cele doua celule-fiice rezultate prin aceasta
diviziune.(fig.11)

Schema structurii unui cromozom la
eucariote (fig 11) Cromatide Brat lung Centromer (constructie primara) Brat scurt
![]()
![]()
![]()
![]()
![]()
![]()
![]()
Dimensiunea cromozomilor. Cromozomii eucariotelor au dimensiuni cuprinse intre 0,2 si 50m in lungime si 0,2- 2m in diametru. De pilda, la ceapa, lungimea cromozomilor este cuprinsa intre 10-20m,la porumb intre8-10m, in timp ce cromozomii umani au intre 2-8m
Forma cromozomilor depinde in primul rand de pozitia centromerului, structura cu ajutorul careia cromozomul se ataseaza de fibra fusului nuclear si are posibilitatea sa migreze la polii celulei in timpul anafazei.
La eucariote, cromozomii au o arhitectura foarte complexa, fiind alcatuiti din 13-15% AND ,12-13% ARN cromozomial, 68-72% proteine histinice si nonhistonice, mici cantitati de lipide si ioni de Mg si Ca.
Cromozomii de tip eucariot sunt alcatuiti din cromatina, care este de doua feluri; eucromatina, substanta cromozomiala care se coloreaza normal si care se replica de-a lungul fazei S a ciclului mitotic, si heterocromatina, mai condensata, care se coloreaza mai intens aproape in toate fazele diviziunii celulare si se replica la sfarsitul fazei S a ciclului mitotic
Tipuri de cromozomi:
-Metacentric cu centromerul median
-Submetacentric cu centromerul plasat submedian
-Subtelocentric cu centromerul plasat aproape de unul din capetele cromozomului
-Acrocentric cu centromerul plasat aproape terminal